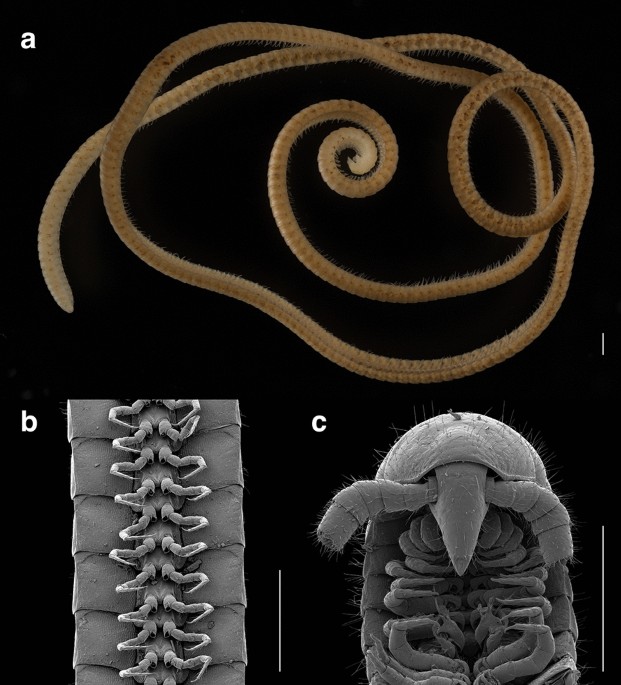
The first true millipede—1306 legs long The name ‘millipede’ translates to a thousand feet (from mille ‘thousand’ and pes ‘foot’). However, no millipede has ever been described with more than 750 legs. We discovered a new record-setting species of millipede with 1,306 legs, Eumillipes persephone, from Western Australia. This diminutive animal (0.95 mm wide, 95.7 mm long) has 330 segments, a cone-shaped head with enormous antennae, and a beak for feeding. A distant relative of the previous record holder, Illacme plenipes from California, it belongs to a different order, the Polyzoniida. Discovered 60 m below ground in a drill hole created for mineral exploration, E. persephone possesses troglomorphic features; it lacks eyes and pigmentation, and it has a greatly elongated body—features that stand in stark contrast to its closest surface-dwelling relatives in Australia and all other members of its order. Using phylogenomics, we found that super-elongation (> 180 segments) evolved repeatedly in the millipede class Diplopoda. The striking morphological similarity between E. persephone and I. plenipes is a result of convergent evolution, probably for locomotion in similar soil habitats. Discovered in the resource-rich Goldfields-Esperance region and threatened by encroaching surface mining, documentation of this species and conservation of its habitat are of critical importance. Among the earliest animals to breathe atmospheric oxygen1 and with some extinct species that grew to two meters in length2, millipedes have lived on this planet for more than 400 million years. Important as decomposers in terrestrial ecosystems3, primary knowledge of millipede diversity lags tremendously behind other animal groups. With fossil taxa dated to the Cretaceous4,5, the millipede order Polyzoniida includes ca. 70 species with a distribution on all continents except Antarctica. Some of its members exhibit parental care of eggs6, others ooze chemical defenses containing alkaloids7 that are sequestered by poison frogs8,9, and some species roll into a ball for protection10. Hatchlings emerge from the egg with four pairs of legs, and continuously add segments during development for an indeterminate period of time, even after adulthood11. Polyzoniidans have never before been observed in deep soil habitats, and most occur in surficial microhabitats of decaying wood and other detritus. Previously known Australian polyzoniidans have at most 400 legs, eyes with 1–3 ommatidia, most with at least some dark pigmentation, a flat wide body (relative to others in the order), and occurrence in epigean microhabitats12. However, distantly related millipedes, such as Illacme species and other members of the family Siphonorhinidae (order Siphonophorida), have been recorded from as deep as 11.5 m below the soil surface13,14. These species possess a suite of adaptations, such as a thin elongated body with up to 750 legs, no eyes, massive antennae, and lack of pigmentation. No siphonorhinid species have been recorded from Australia, and the geographically closest siphonorhinids occur in Madagascar and Indonesia13,14.Here we report the discovery of E. persephone, the first super-elongated millipede known from Australia, and the new world record holder of the animal with greatest number of legs. It belongs to the family Siphonotidae (order Polyzoniida), yet appears similar to super-elongated millipedes in the order Siphonophorida. Eumillipes persephone lives deep underground, and it was only discovered by surveying geological drill holes originally created for mineral exploration that provided access to a cryptic and previously unexplored underground habitat.In August 2020, in the Goldfields region of Western Australia, we discovered a pale, thread-like millipede with 1,306 legs at a depth of 60 m in a drill hole created for mineral exploration (Fig. 1). In total, 56 4–81 m deep 150 mm-diameter drill holes located at a site 100 km WSW of Norseman, Western Australia (within the Great Western Woodlands) were sampled for millipedes and other subterranean fauna, according to techniques described in ref.16. These geological drill holes provided a portal to access an interstitial habitat composed of banded iron formations and mafic volcanic rocks, which are known to harbor troglophilic fauna16,17. Eight individuals of the new species were collected from troglofauna traps at depths between 15–60 m from three drill holes (450 m apart), including five individuals from a trap set at a depth of 60 m. Two juveniles were collected in April 2020 and another one in January 2021, always from troglofauna traps, attesting to the species’ true deep subterranean life.Figure 1The leggiest animal on the planet, Eumillipes persephone, from Australia. (A) female with 330 segments and 1,306 legs (paratype specimen, T147124). (B) ventral view of legs (male holotype, T147101). (C) dorsal view of head and ventral view of gonopods (male holotype, T147101). Scale bars, 0.5 mm.The specimens superficially appeared to be members of the Siphonorhinidae, due to phenotypic similarity to Illacme plenipes Cook & Loomis, 1928. We used scanning electron microscopy and genome sequencing to conclude that the remarkable anatomical similarity between E. persephone and I. plenipes is a result of convergent evolution of super-elongation (> 180 segments), which we here reveal that has originated at least twice in the millipede class Diplopoda (Fig. 2). Eumillipes persephone appears unlike any other species in the order Polyzoniida. Its highly elongated and narrowed body, lack of eyes, massive antennae, shortened legs, and lack of pigmentation comprise a suite of characters consistent with a fossorial bauplan that has repeatedly evolved in the Diplopoda18,19.Figure 2Phylogenomic estimation of the evolutionary history of colobognath millipedes. Super-elongation (> 180 segments) is well known from the order Siphonophorida, including Illacme plenipes with 192 segments and 750 legs, but it has independently evolved in the Australian order Polyzoniida with Eumillipes persephone bearing up to 1,306 legs and 330 segments. This feature (dark branches) has evolved between two and four times in diplopods based on a character state reconstruction using parsimony. Maximum likelihood phylogeny of 312 orthologous sequences with Polydesmida, Stemmiulida, Chordeumatida, Spirostreptida, and Sphaerotheriida as outgroup taxa. Julida, Spirobolida, and other diplopod orders omitted from the analysis. Support values on nodes are bootstrap supports. Numbers in parentheses after species names are the maximum segment count for the taxon. Species with super-elongation present denoted in bold. Millipede silhouettes sized relative to one other. Diagram created with Adobe Illustrator 2021 (adobe.com/products/illustrator.html).Taxonomy sectionClass Diplopoda de Blainville in Gervais, 1844Subterclass Colobognatha Brandt, 1834Order Polyzoniida Cook, 1895Family Siphonotidae Cook, 1895Tribe Rhinotini Hoffman, 1977 Eumillipes, Marek new genushttp://zoobank.org/NomenclaturalActs/a258f64e-0d4c-4941-be20-0b5d78e1078e.Type speciesEumillipes persephone Marek, new species.Generic placement and diagnosisThe genus Eumillipes is placed in the order Polyzoniida, family Siphonotidae, tribe Rhinotini based on the following characters10,20; whether the character is diagnostic for the order (O), family (F), or tribe (T) is denoted in parentheses. Head capsule small, conical, elongated into a sharp snout (O) (Fig. 1C, Supplementary Fig. S1A–C). Prozonites and metazonites of trunk rings the same width (O) (Supplementary Fig. S2A); prozonites not narrow as in the Siphonophorida. Rings smooth (Supplementary Fig. S2B), covered with neither cuticular ornaments nor long setae as in the Siphonophorida. Vertex of head with two macrosetae (O) (Supplementary Fig. S1C). Antennae thick with equally sized antennomeres (O) (Fig. 1C, Supplementary Fig. S3A); antennae not strongly elbowed between antennomeres 3, 4 as in the Siphonorhinidae (Siphonophorida). Gnathochilarium reduced to three sclerites: mentum; left, right stipes (O). Ozopores located far in from lateral edges of tergites; placed two-thirds the distance from midline laterally to tergal margins (F) (Supplementary Fig. S2D). Vasa deferentia open through small conical lobes (penes) on the posterior surfaces of the second leg coxae (F). Telson a complete ring around the anal valves (F) (Supplementary Fig. S2D). Tarsal claw with long sigmoid-shaped accessory claw at its base that exceeds the claw in length (F) (Supplementary Fig. S1D). The genus differs from other siphonotid genera by the following characters. Anterior gonopods (i.e., 9th leg pair) strongly modified with podomeres 3–4 fused, not leg-like as in Siphonotini genera (T) (Fig. 1C, Supplementary figs. S4, S5A). Apex of anterior gonopods distinctly bifurcated into two processes (Fig. 1C, Supplementary figs. S4, S5A), not a single process as in the genera Rhinotus Cook, 1896 and Siphonoconus Attems, 1930. Eumillipes persephone Marek, new species http://zoobank.org/NomenclaturalActs/0AFB7037-E517-4D05-804B-D9AE1C7B3F47 DiagnosisAdults of Eumillipes persephone can be differentiated from other polyzoniidan genera and species (and commonly encountered millipedes co-occurring with E. persephone in Western Australia) based on the combination of the following characters. Body extremely long and thread-like (width: ♂ 0.92 mm; ♀ 0.95 mm; length: ♂ 54.7 mm; ♀ 95.7 mm). Exoskeleton uniformly pale, cream-colored (Fig. 1A, Supplementary figs. S6–S8)—with neither dark pigmentation nor longitudinal, nor transverse stripes as in surface-dwelling species (Supplementary Fig. S9). Adult millipedes with an exceptional number of rings and legs: ♀ up to 330 rings and 1,306 legs, and ♂ up to 208 rings and 818 legs (Supplementary tab. S1). Head cone-shaped and eyeless (Fig. 1C, Supplementary Fig. S1A–C)—lacking eyes as are present in surface-dwelling species. Ninth and 10th leg pairs modified into gonopods (Fig. 1C, Supplementary figs. S4, S5). Anterior gonopods (9th leg pair) highly modified, not leg like, and distinctly bifurcated into two processes (Fig. 1C, Supplementary figs. S4, S5A). Medial process of the anterior gonopods saddle-shaped—not pointed and recurved as in Siphonotus flavomarginatus Attems, 191121. Lateral process sheath-like and cupping the medial process. Posterior gonopods (10th leg pair) stylet-like, and in repose threaded through the bifurcated anterior gonopods (Fig. 1C, Supplementary figs. S4, S5B). Sterna of gonopods with four long, slender, curved setae apically studded with spinules (Supplementary figs. S4B, S5A).Material examinedMale holotype (WAM T147101), two female and one male paratypes (WAM T147100, T147124, T146684), and two juveniles (WAM T147122, T147123) from Western Australia, ca. 100 km WSW of Norseman, 32° 32′ 05.9″ S, 120° 47′ 42.74″ E, 27 May–4 August 2020, collected by A.J. Mittra and L.P. Masarei (Western Australian Museum).VariationMales have fewer segments and legs; specimen T147101 has 198 segments and 778 legs (Supplementary Fig. S7), and T147100 has 208 segments and 818 legs (Supplementary Fig. S8). Female specimen T147124 possesses 330 segments and 1,306 legs (Fig. 1A, Supplementary Fig. S6), and T146684 has 253 segments and 998 legs.EtymologyThe genus is named because it is the first true millipede with more than 1000 legs. The name Eumillipes is a combination of the Greek eu-, meaning ‘true’; Latin mille, ‘thousand’; and Latin pes, ‘foot’. It is to be treated as a noun. The species epithet is derived from the Greek mythological goddess of the underworld, Persephone, who was originally from the surface but was taken to the underworld by Hades.From studies by Manton22 and video footage of siphonorhinid millipedes, the locomotive biomechanics of super-elongated millipedes has only been studied rudimentarily. Navigating by its enlarged antennae (Fig. 1C, Supplementary Fig. S3A, B) equipped with up to five sensillum types15, the millipede’s elongated trunk with telescopic segments facilitates burrowing in a three-dimensional soil matrix. Its antennae wave independently to locate an opening; once a suitable gap is identified, the millipede enters and walks through. The flexible segments are compressible to squeeze through narrow fissures. The continuous metachronal gait and musculoskeletal action of the segments, which are composed of concentric tubular rings that slide within one another, provide continuous pushing force; in combination with the legs, longitudinal (Supplementary Fig. S10C) and oblique muscles pull the rings together and provide forward locomotion22. This way of burrowing resembles that used by geophilomorph centipedes (another group of greatly elongated myriapods that includes the leggiest centipede, Gonibregmatus plurimipes Chamberlin, 1920 from Fiji with 191 rings and 382 legs) and earthworms. In contrast with millipedes with incompressible rings such as ‘bulldozer’ ecomorphs that walk in a single plane propelled by the additive force of their legs22, colobognath millipedes are able to wind their way through a three-dimensional matrix and can simultaneously walk in up to eight different planes. This telescoping locomotion, by sliding trunk segments (Supplementary Fig. S2A) coupled with the thrust of the legs, propels the animal through a varied and unpredictable underground microhabitat22, and the increase in leg number likely contributes more pushing power to force through small crevices and openings. Super-elongation in the Diplopoda has repeatedly evolved in taxa that live in these microhabitats, such as in Siphonorhinidae (I. plenipes with 192 segments and Nematozonium filum Verhoeff, 1939 with 182 segments), Siphonophoridae (Siphonophora millepeda Loomis, 1934 and Siphonacme lyttoni Cook & Loomis, 1928 each with 190 segments), and E. persephone. Each additional ring provides extra locomotory thrust23, and E. persephone’s up to 330 rings may be specially adapted to locomotion in its comparatively deep soil microhabitat at 60 m—five-fold greater than the maximum depth of Illacme species. Trunk super-elongation may also serve to lengthen the digestive canal to increase the absorptive surface area and assimilation efficiency in a resource-limited subterranean habitat, as may be the case for I. plenipes, which additionally has an even longer corkscrew-shaped gut than the trunk itself15. The diet of E. persephone is unknown, but may be similar to other primarily fungivorous colobognath millipedes, such as the platydesmidan Brachycybe lecontii Wood, 1864 that feeds on up to 176 different genera of fungi24.Current alpha-level knowledge of Australian Siphonotidae is fragmentary. Although an unpublished dissertation from 1994 documented eight genera and 42 species of the family25, taxonomy of the group remains antiquated with two genera and five species currently listed from the continent described about a century ago21,25,26. Although the phenotypic divergence of E. persephone from surface-dwelling family relatives appears to be associated with adaptations to life underground, extraordinary similarity of its gonopods (species-diagnostic features in diplopods) with Siphonotus flavomarginatus from Torbay, Western Australia, indicates a genus-level affiliation. Close similarity of gonopod anatomy with extremely divergent somatic features suggests rapid evolution of troglomorphic traits.Invertebrates tha
https://www.nature.com/articles/s41598-021-02447-0
The first true millipede—1306 legs long