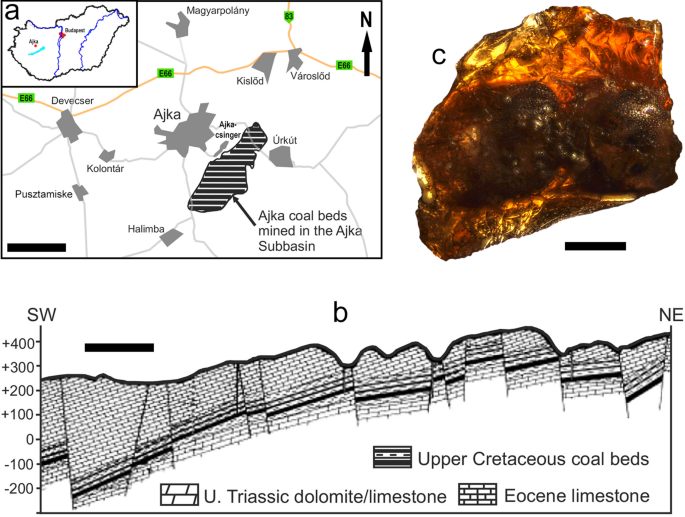
The first mainland European Mesozoic click-beetle (Coleoptera: Elateridae) revealed by X-ray micro-computed tomography scanning of an Upper Cretaceous amber from Hungary Scientific Reports 12, Article number: 24 (2022) Cite this article EntomologyEvolutionPalaeontologyTaxonomyZoology Fossil bioinclusions in amber are invaluable source of information on the past evolution and diversity of various organisms, as well as on the paleoecosystems in general. The click-beetles, Elateridae, which originated and greatly diversified during the Mesozoic, are mostly known from the adpression-like fossils, and their diversity in the Cretaceous ambers is only poorly documented. In this study, we describe a new click-beetle based on an incomplete inclusion in ajkaite, an Upper Cretaceous (Santonian) amber from the Ajka Coal Formation from Hungary. We used X-ray micro-computed tomography scanning to reconstruct its morphology because it is deposited in an opaque piece of amber. Our results suggest that the newly described Ajkaelater merkli gen. et sp. nov. belongs to subfamily Elaterinae. It represents the first Mesozoic beetle reported from Hungary, and the first Mesozoic Elateridae formally described from mainland Europe. Our discovery supports an Eurasian distribution and diversification of Elaterinae already in the Cretaceous. The paleoenvironment of the Ajka Coal Formation agrees well with the presumed habitat preference of the new fossil taxon. The discovery of a presumably saproxylic click-beetle shed further light on the yet poorly known paleoecosystem of the Santonian present-day western Hungary. Fossils play an important role in our understanding of past processes including the origin, evolution and diversification of beetle lineages1,2. Among fossils, amber inclusions are especially valuable for scientists because they allow to observe the morphological features in much better detail than e.g., compression fossils3,4. Mesozoic amber outcrops are known from various places on the planet5,6 but only a few of them dated to the Cretaceous include fossil insects. The most important Cretaceous amber sites are located in Lebanon, Israel, Jordan, France, England, Spain, USA, Canada, Russia and Myanmar5,6,7,8,9,10,11,12,13,14,15. The Burmese, Lebanese, Spanish and French sites are particularly interesting due to their great abundance and diversity of insects5,14,15,16. While some amber sites produce many bioinclusions, with numerous new insect taxa being described on a regular basis, some other amber sites are rather poor in the matter of described diversity. For example, bioinclusions have been found only in a fraction of amber sites in France15,17, most probably due to a high proportion of the opaque amber stones, in which the inclusions can be detected only using X-ray radiographic and tomographic imaging techniques15.Upper Cretaceous amber-bearing sedimentary units in Hungary include the alluvial floodplain deposits of the Csehbánya Formation nearby Iharkút and the Ajka Coal Formation (Ajkacsinger) southeast from the city Ajka in the Bakony Mountains in southwestern Hungary (Fig. 1), both being Santonian in age. Despite more than a thousand of amber pieces known from the former locality, no inclusions have been found in these dominantly small (up to 3 mm), drop-like pieces so far. On the other hand, the unique type of amber from the brown coal beds of Ajka, so-called ajkaite18, contains many bioinclusions.Figure 1Ajka location and geology, and the here examined amber inclusion. (a) Location of Ajka in western Hungary, with the location of the Ajka coal beds. (b) Simplified geological section of the Ajka Subbasin. (c) Ajkaite specimen containing the holotype of Ajkaelater merkli gen. et sp. nov., under polarized light microscope. Scale bars: (a) 3 km; (b) 1 km; (c) 1 mm.The Ajka Coal Formation, being at some places over 100 m thick19,20, comprises an alternation of coal beds, carbonaceous to argillaceous pelitic sediments with interbedded molluscan lumachelles, marls, and sandstone beds representing a lacustrine-palustrine sequence21. It has a well documented fossil flora22,23,24,25,26, and fauna of mollusks27,28 and vertebrates29. Despite a relatively large number of available amber stones from the Ajka deposits, the record of the ajkaite inclusions is still rather scarce. Short mentions on the inclusions in ajkaite are known since the middle of the twentieth century30,31. Regarding the arthropods, apart from an officially unpublished MSc thesis32, there is only one detailed study by Borkent33 describing two species of ceratopogonid flies. Obviously, many other ajkaite arthropods belonging to Arachnida, Diptera, Hymenoptera and Coleoptera are waiting to be formally described34.The Elateridae, commonly known as click beetles, are the largest and most diverse family in the superfamily Elateroidea. Adult individuals can be usually recognized by their elongate, narrow body, structure of basal antennomeres, pro-mesothoracic clicking mechanism, acute posterior angles of pronotum, and five abdominal ventrites, of which four are connate and the last one is free35,36,37. Their most characteristic feature is their ability to jump into the air by rapidly sliding their prosternal process into their mesosternal cavity, with a typical clicking sound, giving their common name35,37. The family currently includes more than 11,000 described species worldwide37,38 and the fossil record includes 261 species in 99 genera39,40. Although they are quite common in various amber deposits worldwide, only a few of them have been described so far40. Regarding the Cretaceous click-beetles in amber, only three species have been described to date, all from the Burmese amber41,42.In this study, we describe a new click-beetle inclusion in ajkaite, which represents the first Mesozoic beetle reported from Hungary as well as the first Mesozoic Elateridae described from the mainland Europe. Since this incomplete specimen is deposited in a non-transparent piece of amber, we had to use X-ray micro-computed tomography scanning to reveal its morphology. We discuss the probable ecology of a new click-beetle in connection with the Ajka Coal paleoenvironment.We describe here Ajkaelater merkli gen. et sp. nov. (Figs. 2, 3, 4, 5, Supplementary File 1) from Santonian deposits of Ajka based on an incomplete specimen embedded in amber. Based on its habitus and morphology of prothorax, we place it into the click-beetle subfamily Elaterinae, without a tribal assignment (see ‘Discussion’ section).
Systematic paleontology Family Elateridae Leach, 1815Subfamily Elaterinae Leach, 1815Tribe Incertae sedis Ajkaelater gen. nov. (Figs. 2, 3, 4, 5, Supplementary File 1)urn:lsid:zoobank.org:act:F3771C8D-2247-4967-A588-B5505E41AB97Figure 2Ajkaelater merkli gen. et sp. nov., holotype (MTM PAL 2021.50.1.). (a) Dorsal view; (b) ventral view; (c) right lateral view. Scale bar: 1 mm.Figure 3Ajkaelater merkli gen. et sp. nov., holotype (MTM PAL 2021.50.1.). (a) Preserved antennomeres of the right antenna (in right oblique ventral view); (b) prothorax in dorsal view; (c) prothorax in ventral view; (d) prothorax in left oblique ventral view. Scale bars: 1 mm.Figure 4Ajkaelater merkli gen. et sp. nov., holotype (MTM PAL 2021.50.1.). Cross section along the saggital axis. e elytron, mv metaventrite, s scutellum, pn pronotum, ps prosternum, psp prosternal process. Scale bar: 1 mm.Figure 5Artistic reconstruction of Ajkaelater merkli gen. et sp. nov. in the swampy Ajka coal area in the Santonian western Hungary, with Hungarosaurus Ősi, 2005 in the background.Type speciesAjkaelater merkli gen. et sp. nov.; by present designation.DiagnosisAdult. Body small, oblong-ovate, presumably about 3.25 times as long as wide. Antenna slightly serrate, presumed antennomeres VI–X about twice as long as wide. Pronotum slightly wider than long when measured along midline, widest at posterior angles; lateral sides weakly rounded; posterior angles sharp, slightly divergent; sublateral carina distinct, short; lateral carina distinct, complete; both lateral and sublateral carinae divergent anterad in lateral view. Pronotosternal sutures slightly convergent posterad, then distinctly curved near procoxal cavities. Prosternum elongate and rather narrow, including prosternal process about twice as long as wide, widest near procoxal cavities; prosternal process relatively long, subparallel-sided, abruptly narrowed before apex in lateral view so that apex is on different plane than rest of prosternum. Scutellar shield longer than wide, with anterior margin slightly rounded and steeply declivous. Mesoventral cavity with sides subparallel-sided for greater part of their length, narrowing slightly between mesocoxae. Elytra oblong-ovate, slightly wider than prothorax, sides slightly rounded; each elytron with distinct impressed striae formed by lines of large punctures. For more details, see the description of A. merkli gen. et sp. nov. below, Figs. 2, 3, 4, 5, and Supplementary File 1.EtymologyDerived from the words ‘Ajka’ (referring to the city of Ajka, the geographic origin of the fossil) and ‘Elater’ (a genus name in Elateridae). Gender: masculine.Composition and distributionOnly A. merkli gen. et sp. nov. (Santonian of Hungary). Ajkaelater merkli gen. et sp. nov. (Figs. 2, 3, 4, 5, Supplementary File 1)urn:lsid:zoobank.org:act:15B434D2-B294-4021-B39F-3AE53A846464Type materialHolotype, adult, sex unknown, inventory number MTM PAL 2021.50.1. (MTM). A beetle fragment is included in a dark reddish amber piece with dimensions of approximately 4.4 × 3.8 × 1.1 mm (Fig. 1c). There are no visible syninclusions in the amber stone.Type horizon and age. Ajka Coal Formation, unknown shaft of the Ajka-Csingervölgy coal minery. Based on palynological and nannoplankton investigations, the age of the formation is dated as Upper Cretaceous (Santonian; 86.3–83.6 Ma)22,29,43,44.Type locality. Ajka-Csingervölgy [Ajka-Csinger valley], approximately 1 km SE of the city Ajka, Bakony Mountains, Hungary (Fig. 1a).DescriptionBody (Figs. 2, 3, 5) about 6.50 mm long (rough guess), 2.00 mm wide (measured at humeri), oblong-ovate, subparallel-sided, slightly convex. Head missing, only parts of right antenna preserved, probably with antennomeres VI–IX and X–XI. Preserved antennomeres (Figs. 2b, 3a) weakly serrate, presumed antennomeres VI–X subequal in length, about twice as long as wide, apical antennomere simple, about 1.20 times as long as penultimate antennomere. Pronotum (Figs. 2a, 3b) slightly wider than long when measured along midline (length: 1.75 mm, width: 1.90 mm), widest at posterior angles but only slightly wider there than in about middle. Anterior angles inconspicuous; lateral sides from dorsal view weakly rounded; posterior angles moderately long, sharp, slightly divergent, each medially with distinct sublateral carina running from apex of angle and slightly surpassing posterior margin of pronotum, not subparallel with lateral carina but both carinae divergent anterad in lateral view (Fig. 2c); posterior margin medially with shallow arcuate indentation. Lateral carina (Fig. 2c) distinct, complete, very weakly and gradually sinuate near middle and slightly more sinuate near posterior angle in lateral view. Disc (Figs. 2a, 3b) moderately convex, relatively densely covered with rounded shallow punctures (although punctures not well visible on most micro-CT scans and if visible, then usually only near margins). Hypomeron mostly smooth, with small punctures. Pronotosternal sutures (Figs. 2b, 3c,d) slightly convergent posterad, then distinctly curved near procoxal cavities, their anterior part not well preserved but most probably slightly excavate anteriorly (Fig. 3d). Prosternum (Figs. 2b, 3c,d) elongate, including prosternal process about twice as long as wide, in front of procoxal cavities (i.e., excluding prosternal process) about 1.30 times as long as wide, widest near procoxal cavities; anterior part not well preserved. Prosternal process (Fig. 3d) relatively long, about 0.55 times as long as prosternum in front of procoxal cavities, subparallel-sided, abruptly narrowed before apex in lateral view so that apex is on different plane than rest of prosternum (Fig. 4); apex narrowly rounded. Procoxal cavities subcircular, moderately widely separated by width of prosternal process. Scutellar shield (Figs. 2a, 3b) about 1.50 times as long as wide, anterior margin well defined, angulate, slightly rounded and steeply declivous; lateral sides sinuate, narrowed before middle and widest after middle; apex rounded. Mesoventral cavity deep, with well-defined walls; sides subparallel for greater part of their length, narrowing slightly between mesocoxae. Mesocoxal cavities narrowly separated. Elytra (Fig. 2a) oblong-ovate, slightly wider than prothorax, sides slightly rounded; each elytron with nine distinct impressed striae formed by lines of large subcircular punctures, which are separated by about diameter of a puncture; interstriae flattened, smooth. Epipleuron (Fig. 2c) well developed, wide basally, then gradually narrowed near metacoxae. Fore leg (Fig. 2b) slender, moderately long; profemur about as long as protibia, protarsomere I elongate, about 3 times as long as wide, remaining tarsomeres not preserved; remaining legs incomplete or not preserved. Immature stages unknown.EtymologyThe specific name ‘merkli’ is a patronym in honor of late Dr. Ottó Merkl (1957–2021), one of the most prominent Hungarian entomologists, the World expert in darkling beetles (Tenebrionidae), and the long-term curator of the Coleoptera Department of the Hungarian Natural History Museum.Recent rise of interest in research of amber fossils significantly increased the rate of discoveries of various interesting animal lineages with great importance for the understanding of their origin, evolution, and paleodiversity, as well as of the composition of past ecosystems4. The here reported discovery of Ajkaelater merkli gen. et sp. nov. in the Upper Cretaceous Hungarian amber is of great importance for several reasons.It is not only the first formally described beetle from ajkaite, but also the first fossil beetle described from the present-day Hungary. So far, some yet undescribed fossil beetles were found in the Miocene and Pliocene deposits of Hungary. The representatives of Dytiscidae and Staphylinidae were reported from the Late Miocene (Sarmatian) of the Tokaj Mountains45. Dystiscidae, Carabidae Cerambycidae, Cantharidae, Chrysomelidae, and various Curculionoidea were identified in the Pliocene alginite deposits of Pula in Bakony Mountains46,47. Additionally, Krzemiński et al.46 reported Carabidae, Hydrophilidae, Silphidae, Chrysomelidae, Lagriinae, and Apionin
https://www.nature.com/articles/s41598-021-03573-5