The evolution of MRSA demonstrates its genetic adaptation into a first-class multidrug-resistant pathogen. Following the introduction of penicillin and methicillin, S. aureus rapidly developed resistance to these β-lactam compounds15. In the 1950s, vancomycin was used to treat MRSA infections, but recently the emergence of MRSA strains that are less sensitive to vancomycin (VISA) or even resistant to vancomycin (VRSA) means that clinicians are less confident when dealing with MRSA. Therefore, there is an urgent requirement to explore effective drugs for MRSA16. Plants that grow in the natural environment are affected by pathogenic microorganisms, such as bacteria and viruses. Some plants have developed two major strategies to defend against pathogenic microorganisms: (1) plants have a lot of epidermal hairs, a thick waxy stratum corneum, and small stomata to block the invasion of pathogenic microorganisms; and (2) plants produce antibiotic substances, such as tannins and alkaloids to inhibit or kill invading pathogenic microorganisms17. Therefore, potential antimicrobial chemicals could be found in plants to treat pathogenic microorganisms, such as bacteria.
Salvia miltiorrhiza (Danshen) has been widely used in traditional Chinese medicine to treat a variety of diseases, which include coronary artery disease, acute ischemic stroke, hyperlipidemia, chronic renal failure, chronic hepatitis, and Alzheimer’s disease. In addition, Danshen has no serious adverse effects. The main bioactive constituents of S. miltiorrhiza include water-soluble phenolic acids and lipophilic diterpenoid tanshinones18. CT is one of the major tanshinones that were isolated from the roots of the Danshen plant. Recent studies have shown that CT has the potential to treat and prevent the previously mentioned diseases and it is a potent antibiotic agent19.
In this study, the antibacterial effect of CT on MRSA was tested in vitro, and the MICs of CT on MRSA reference strain ATCC43300 and seven clinical strains were detected using a two-fold serial dilution method. The results showed that CT displayed bacteriostatic action against MRSA, which was consistent with previous reports14, which suggests that CT could be used to fight MRSA infection.
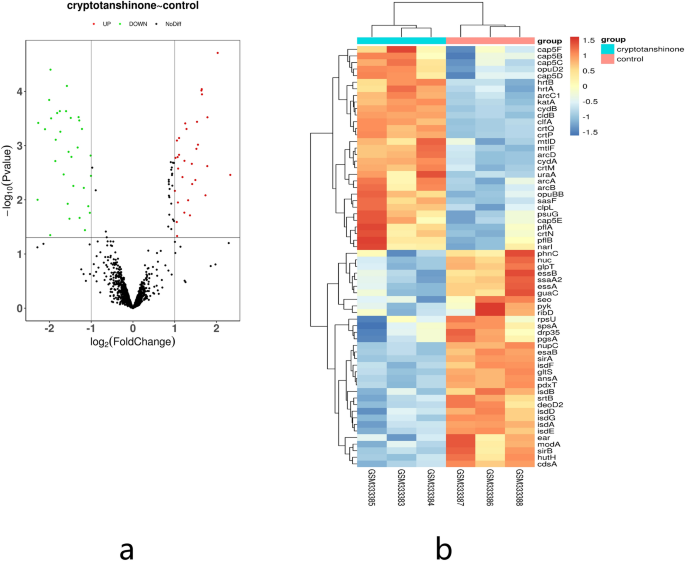
Although thousands of herbal compounds have been listed as antimicrobial phytochemicals20, limited understanding of the mechanisms limits the application of these substances. To determine the mechanism of CT inhibition on the growth of MRSA, bioinformatics tools were used. In this study, the data from was retrieved from GSE13203 and systematically analyzed the differential gene expression of S. aureus were systematically analyzed between the CT treatment and control. Therefore, 64 DEGs were identified, which included 33 downregulated and 31 upregulated genes.
Then, GO and KEGG analysis of these DEGs revealed that CT had a greater impact on the metabolism of S. aureus. Because MRSA is a special species of S. aureus that carries a multidrug resistance gene, the metabolic pathways of MRSA and S. aureus are similar. As a facultative anaerobe, S. aureus uptakes a variety of nutrients that include glucose, mannose, mannitol, glucosamine, N-acetylglucosamine, sucrose, lactose, galactose, and beta-glucosides. The central pathways for glucose metabolism are the Embden–Meyerhof–Parnas pathway and the pentose phosphate cycle. Lactate is the end product of anaerobic glucose metabolism and acetate, and CO 2 are the products of aerobic growth conditions21. In the glucose metabolic pathway, PK is the rate-limiting enzyme of energy metabolism, which plays a central role in the carbohydrate metabolism of MRSA. It catalyzes the final rate-limiting step of glycolysis. During this irreversible process, the high-energy phosphate bond in the PEP molecule is transferred to ADP to generate ATP. The metabolites PEP and pyruvate are related to other biosynthetic pathways, and therefore, the potential inhibition of PK could obstruct MRSA energy metabolism. The potential inhibition of PK might result in the decreased metabolism of MRSA13,22,23,24.
A PK test kit was used to detect the effects of CT on the activity of PK of MRSA in vitro. The results showed that except for in the MRSA2 and MRSA5 groups, CT hampered the PK activity of MRSA. PK was identified as a highly interconnected essential hub protein in MRSA, with structural features distinct from human homologs. Currently, the majority of antibiotics in use are directed at critical proteins that are unique to the bacteria and without human homologs to avoid mechanism based toxicity. In addition, based on the supposition that hub proteins are critical for bacterial survival and they should be very sensitive to mutations25, and therefore, targeting them should reduce the potential to develop resistant strains and species. Therefore, CT could be a valuable antibacterial candidate because of its effect on the PK of MRSA.
In this study, CT did not affect PK activity in MRSA2 and MRSA5, which were collected from a blood specimen and the five other strains were isolated from a sputum specimen. In different environments, S. aureus will utilize a variety of metabolic pathways, which depend on the oxygen content, source of carbohydrates, and other factors26. However, the MIC of CT was 0.9 μg/mL for MRSA2, and 1.9 μg/mL for MRSA5, which indicated that CT might have additional mechanisms that affect the growth of MRSA in addition to affecting the PK activity of both strains. As shown in Table 2, in addition to the pyk gene, a significant downregulation of isdB and isdG were observed, both of which belong to the iron-regulated surface determinant family. The GO-MF analyses revealed that CT affected the metal ion binding of S. aureus. Iron is vital for the growth and proliferation of nearly all organisms, including MRSA. In addition, iron is required for the colonization of host tissues by MRSA and subsequent pathogenesis27. Therefore, in addition to affecting PK activity, CT might inhibit MRSA growth by interfering with its iron metabolism, however, this requires further investigation.
Table 2 Top10 up and downregulated genes in DEGs. Full size table
The most significant limitation of this study was that only changes in the activities of PK MRSA were examined after treatment with CT. Future research should include measuring the expression of PK of MRSA after treatment with CT. In addition, only seven MRSA strains were isolated from a clinical environment; therefore, a larger number of samples is required in future research to verify the antimicrobial mechanism of CT on MRSA.
In summary, CT inhibited the growth of MRSA in vitro. To determine the mechanism of activity, multiple bioinformatics tools were used combined with a comprehensive analysis of gene expression profiles to identify the key signaling pathways. CT interfered with the activities of PK, which is the key rate-limiting enzyme during glycolysis in MRSA. The results of this study might provide new insights into the antibacterial mechanisms of CT. However, further research and studies with larger sample sizes are required to confirm these findings.
http://feeds.nature.com/~r/srep/rss/current/~3/0R9NrOeMqYc/s41598-021-01121-9