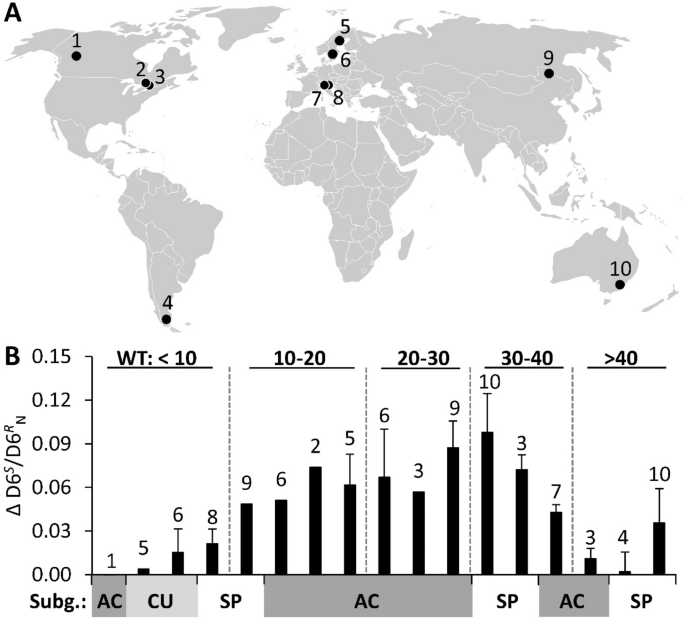
Global CO2 fertilization of Sphagnum peat mosses via suppression of photorespiration during the twentieth century Henrik Serk1,2, Mats B. Nilsson2, Elisabet Bohlin2, Ina Ehlers1, Thomas Wieloch1, Carolina Olid2,3, Samantha Grover4, Karsten Kalbitz5, Juul Limpens6, Tim Moore7, Wiebke Münchberger8, Julie Talbot9, Xianwei Wang10, Klaus-Holger Knorr8, Verónica Pancotto11 & Jürgen Schleucher1 Scientific Reports 11, Article number: 24517 (2021) Cite this article Natural peatlands contribute significantly to global carbon sequestration and storage of biomass, most of which derives from Sphagnum peat mosses. Atmospheric CO2 levels have increased dramatically during the twentieth century, from 280 to > 400 ppm, which has affected plant carbon dynamics. Net carbon assimilation is strongly reduced by photorespiration, a process that depends on the CO2 to O2 ratio. Here we investigate the response of the photorespiration to photosynthesis ratio in Sphagnum mosses to recent CO2 increases by comparing deuterium isotopomers of historical and contemporary Sphagnum tissues collected from 36 peat cores from five continents. Rising CO2 levels generally suppressed photorespiration relative to photosynthesis but the magnitude of suppression depended on the current water table depth. By estimating the changes in water table depth, temperature, and precipitation during the twentieth century, we excluded potential effects of these climate parameters on the observed isotopomer responses. Further, we showed that the photorespiration to photosynthesis ratio varied between Sphagnum subgenera, indicating differences in their photosynthetic capacity. The global suppression of photorespiration in Sphagnum suggests an increased net primary production potential in response to the ongoing rise in atmospheric CO2, in particular for mire structures with intermediate water table depths. Over one third of global soil carbon (C) is stored in boreal mires1,2, making peat C accumulation an essential part of the global C budget. Changes in climate are expected to have strong effects on peatland C sequestration1,3,4. During the early and mid-Holocene, the accumulation of peat C was largely determined by the retreat of the northern ice sheet and the rise in temperature because atmospheric CO2 concentrations were relatively stable at 275 ± 8 ppm (SD)5,6,7. Since the beginning of the industrial revolution in the early nineteenth century, CO2 concentrations have risen from ca. 280 ppm to over 400 ppm today8. Multiple observations indicate that recent increases in atmospheric CO2 have affected peat C accumulation rates: (i) the variation in acrotelm peat accumulation was mainly driven by photosynthesis9, (ii) peat C accumulation in Alaskan mires increased about threefold during the twentieth century10, and (iii) the variation in net ecosystem exchange between mires was mainly controlled by differences in leaf area index11. In addition to rising atmospheric CO2 levels, ongoing climatic changes such as increases in temperature and changes in precipitation are hypothesized to influence peatland C fluxes12,13,14.Sphagnum peat mosses are primarily responsible for the accumulation of peat C because they often constitute 80–100% of the ground cover in northern peatlands15. Compared to vascular plants, Sphagnum remnants are highly resistant to microbial decay, which is vital for peat C accumulation16,17. Therefore, C accumulation and storage in the form of Sphagnum remains generally exceeds C losses from microbial decay. However, currently it is not clear whether increases in Sphagnum C accumulation driven by ongoing and projected global warming will outweigh increases in the rate of microbial peat decomposition9,18,19. Understanding how Sphagnum C fluxes respond to recent and projected increases in atmospheric CO2 is therefore crucial for predicting future peat C fluxes.To our knowledge, responses of Sphagnum photosynthetic C fluxes to the recent increase in atmospheric CO2 have never been explored on the global scale. Previous attempts to estimate responses of Sphagnum to increased atmospheric CO2 were either based on free-air CO2 enrichment (FACE) or greenhouse experiments20,21,22,23,24,25. A recently developed isotopomer method26,27 enables reconstruction of metabolic C fluxes by analyzing cell wall carbohydrates from Sphagnum remnants. This approach involves using NMR spectroscopy to measure the abundance ratio of the deuterium (D) isotopomers D6S and D6R in the C6H2 groups of glucose derived from hydrolyzed cell wall carbohydrates, where S and R are the stereochemical designators. The abundance ratio of these D isotopomers is correlated with the ratio of Rubisco oxygenation to carboxylation due to D fractionation in the photorespiration pathway26. Therefore, the D6S/D6R ratio reflects the relative rates of photorespiration and (gross) photosynthesis which essentially depend on the substrate ratio of CO2 and O2. The uptake of C in C3 plants is facilitated by the carboxylation reaction, catalyzed by the enzyme Rubisco. The oxygenation activity of this enzyme, however, leads to the loss of C and therefore reduces the efficiency of photosynthesis. This process is known as photorespiration, which accounts for C losses of up to 25%, and therefore plays an important role for the global terrestrial C sink28.Most studies focus on photorespiration in higher plants and only little is known about Sphagnum mosses, despite their importance for global peatland C fluxes. Using the isotopomer method, we recently experimentally investigated the response of the photorespiration to photosynthesis ratio (i.e. the D6S/D6R ratio) to the recent increase in atmospheric CO2 levels from 280 to 400 ppm and the dependence of this response on selected climate variables including temperature, water table (WT) depth, and light intensity27. We found that under low WT conditions (20 cm below moss surface), photorespiration was suppressed relative to photosynthesis in the hummock species Sphagnum fuscum. Under water-saturating conditions (WT near moss surface), however, there was no effect of atmospheric CO2, indicating that WT depth strongly influences the CO2 fertilization effect in S. fuscum. In addition, the lawn species S. majus did not respond to the CO2 increase, suggesting a species-specific response. Therefore, Sphagnum photosynthetic C fluxes are expected to vary with species and microhabitat29,30,31,32.The aim of this study is to investigate changes in Sphagnum photosynthetic C fluxes during the twentieth century at the global scale. To do this, we estimated the global response of the photorespiration to photosynthesis ratio by comparing the D6S/D6R ratios of modern Sphagnum tissues formed at the current atmospheric CO2 concentration (ca. 400 ppm) to the D6S/D6R ratios of Sphagnum remnants from peat core sections formed at least 100 years ago under pre-industrial CO2 concentrations (≤ 300 ppm). Our analysis is based on 36 peat cores from 10 different sites on five continents (Fig. 1A). Effects of microhabitat were tested by including both hummock (n = 25) and lawn (n = 11) samples with WT depths ranging from ≈ 5 to 70 cm below the moss surface. The effect of species was tested by including eight different Sphagnum species belonging to three different taxonomic sections (subgenera). To determine whether the CO2 response of the photorespiration to photosynthesis (D6S/D6R) ratio in Sphagnum was influenced by changes in temperature and precipitation during the twentieth century, we estimated these changes using established climate models33,34,35. The effects of changes in WT depth during the twentieth century were estimated using WT reconstruction data available in the literature and by measuring δ13C, which has been proposed as proxy for surface moisture27,36.Figure 1Global changes in the deuterium isotopomer ratio (ΔD6S/D6RN) of Sphagnum during the twentieth century representing changes in the photorespiration to photosynthesis ratio. (A) Global distribution of investigated sites. (B) Response of the D6S/D6R ratio per unit change in 1000/[CO2] between modern and historical Sphagnum samples (ΔD6S/D6RN). Five water table depths (WT) categories (in cm) are indicated by vertical dashed lines. Sphagnum subgenera are indicated on the x-axis by grey/white shading: AC, ACUTIFOLIA (dark grey); CU, CUSPIDATA (light grey); SP, SPHAGNUM (white). Error bars indicate standard error, n = 1–4 (see Table S1 for more information). Numbers above error bars correspond to sample sites as numbered in (A).Global changes in the photorespiration to photosynthesis ratio during the twentieth century were assessed by comparing the D6S/D6R ratios of modern and ≥ 100 years old Sphagnum tissues. Modern Sphagnum samples were retrieved from surface peat (top 0–2 cm) formed at contemporary atmospheric CO2 levels (ca. 400 ppm). Conversely, historical Sphagnum tissues were retrieved from peat layers ≥ 30 cm below the surface, with an approximate age of 100 years or more (Table S1). Therefore, the historical Sphagnum tissues were formed when atmospheric CO2 concentrations were ≤ 300 ppm. The D6S/D6R ratio of modern Sphagnum was 0.860 ± 0.004 (average ± SE, range: 0.810–0.927), while that of historical Sphagnum was 0.901 ± 0.005 (average ± SE, range: 0.858–0.971, Fig. S1). Thus, the D6S/D6R ratio of modern Sphagnum was generally lower compared to ≥ 100 years old Sphagnum, indicating that photorespiration is suppressed relative to photosynthesis.The peat samples essentially differed with respect to their depth below the surface (which is related to the atmospheric CO2 concentration when the Sphagnum biomass was formed), the Sphagnum subgenus, the present WT depth, and the geographical location (site). The effects of these factors on the D6S/D6R ratio were tested by a linear mixed effect model with the site as random factor, and atmospheric CO2, WT and subgenus as fixed factors. Analysis of variance (ANOVA) of the random factor revealed that the effect of site was not significant (variance = 0.0, p = 1.0). ANOVA of the fixed factors showed a significant effect for atmospheric CO2, WT and subgenus, explaining 35%, 12% and 8% of the variance, respectively (Table 1). A significant interaction was found between CO2 and WT, which explained 12% of variance, indicating that the effect of CO2 on the D6S/D6R ratio is dependent on the WT depth.Table 1 Summary of ANOVA results for linear mixed effect models of the effects of atmospheric CO2, water table (WT) and Sphagnum subgenus (Subg.) on the D6S/D6R ratio.The effect of atmospheric CO2 on the D6S/D6R ratio clearly depended on the present WT depth. The post-hoc test showed that for peat cores where the WT was 0.05, Fig. S1) between modern and historical Sphagnum, with means of 0.893 ± 0.01 and 0.905 ± 0.009 (SE), respectively. Conversely, for peat cores with WT depths of 10–40 cm, the D6S/D6R ratios of modern and historical Sphagnum clearly differed (p 40 cm, the difference between modern and historical samples was again not significant (p > 0.05), with means of 0.860 ± 0.007 and 0.873 ± 0.004 (average ± SE), respectively.For modern Sphagnum, the effect of WT was evident from generally higher D6S/D6R ratios for WT depths of 10 cm (mean 0.852 ± 0.007 SE; Fig. S1). For ≥ 100 years-old samples, this was not the case, instead the D6S/D6R ratio was generally lower for WT depths > 40 cm (mean 0.873 ± 0.004 SE) compared to WT depths of 0.05) between modern and historical samples (Figs. 1 and S1).The differences in the D6S/D6R ratio between modern and historical Sphagnum were normalized based on the linear relationship between the D6S/D6R ratio and 1000/[CO2] previously reported by Ehlers et al.26, to account for variations in atmospheric CO2 concentrations due to differences in peat depth and/or age. To this end, the regression slope of this linear function was calculated as the change in the D6S/D6R ratio per unit change in 1000/[CO2] between modern and historical Sphagnum samples (denoted ΔD6S/D6RN). ΔD6S/D6RN thus represents the degree of suppression of photorespiration; its mean was 0.044 ± 0.008 (SE) and it varied between 0.000 and 0.094 (Fig. 1). ΔD6S/D6RN varied with the WT depth: it was 0.010 ± 0.005 (average ± SE) for WT depths 40 cm (Fig. 1). These results indicate that a WT between 10 and 40 cm below the moss surface is optimal for suppressing photorespiration in response to increased atmospheric CO2.Effect of changes in water table, temperature and precipitation during the twentieth centuryThe relationship between ΔD6S/D6RN and the present WT depth assumes that hydrological conditions were relatively stable over the twentieth century. To test this assumption, we performed a literature review on changes in WT depth during the twentieth century based on
https://www.nature.com/articles/s41598-021-02953-1
Global CO2 fertilization of Sphagnum peat mosses via suppression of photorespiration during the twentieth century