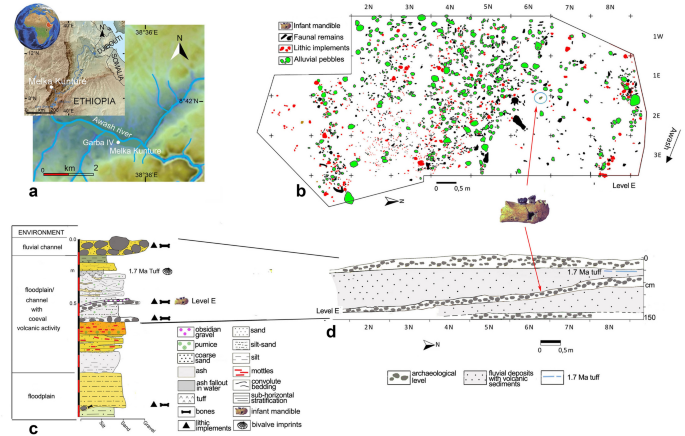
Insights into the palaeobiology of an early Homo infant: multidisciplinary investigation of the GAR IVE hemi-mandible, Melka Kunture, Ethiopia Adeline Le Cabec1,2, Thomas Colard3, Damien Charabidze4, Catherine Chaussain5, Gabriele Di Carlo6, Sabine Gaudzinski-Windheuser7, Jean-Jacques Hublin1, Rita T. Melis8,9, Laura Pioli9, Fernando Ramirez-Rozzi5,10 & Margherita Mussi8,11 Scientific Reports 11, Article number: 23087 (2021) Cite this article ArchaeologyBiological anthropology Childhood is an ontogenetic stage unique to the modern human life history pattern. It enables the still dependent infants to achieve an extended rapid brain growth, slow somatic maturation, while benefitting from provisioning, transitional feeding, and protection from other group members. This tipping point in the evolution of human ontogeny likely emerged from early Homo. The GAR IVE hemi-mandible (1.8 Ma, Melka Kunture, Ethiopia) represents one of the rarely preserved early Homo infants (~ 3 years at death), recovered in a richly documented Oldowan archaeological context. Yet, based on the sole external inspection of its teeth, GAR IVE was diagnosed with a rare genetic disease–amelogenesis imperfecta (AI)–altering enamel. Since it may have impacted the child’s survival, this diagnosis deserves deeper examination. Here, we reassess and refute this diagnosis and all associated interpretations, using an unprecedented multidisciplinary approach combining an in-depth analysis of GAR IVE (synchrotron imaging) and associated fauna. Some of the traits previously considered as diagnostic of AI can be better explained by normal growth or taphonomy, which calls for caution when diagnosing pathologies on fossils. We compare GAR IVE’s dental development to other fossil hominins, and discuss the implications for the emergence of childhood in early Homo. The pattern and timing of modern human life history is unique in involving an extended period of growth1. This provides more time not only for somatic development and protracted brain growth2, 3, but also to learn survival skills4. The offspring is thus dependent upon more parental care over a longer time, and sexual maturity is postponed5. In modern humans, life history stages successively involve infancy, childhood, a juvenile phase, adolescence, and finally adulthood. Different kinds of feeding strategies and dental development stages characterize these phases. Following these criteria, infancy can be subdivided into two stages. First, the ‘nursing phase’ takes place from birth to 6 months, when the infant exclusively relies on breastfeeding and starts erupting its anterior deciduous teeth. Second, the ‘suckling phase’ spans from 6 months to 3 years and involves supplementing complementary food to supply the nutrients not sufficiently present in quantity or quality in the mother milk to enable the fast postnatal brain growth, and the eruption of all deciduous teeth. Weaning is one of the pivotal life history traits occurring during ontogeny: this is a long process involving a transition from exclusive breastfeeding to the progressive introduction of solid food, towards total cessation of mother milk consumption (fully weaned infants) and full reliance on solid diet6, 7, and eventually feeding self-sufficiency. During hominin evolution, major changes in weaning and feeding behaviour may have occurred with early Homo, especially involving the reduction in dental size and dietary shifts to focus on food items more energy dense and easier to masticate, and on an increase in meat and fat consumption and food sharing3, 8, 9. Known as the ‘Expensive-Tissue Hypothesis’8, this energy-rich diet would have enabled meeting the requirements of this demanding postnatal brain growth and achieving adult brain size by the end of childhood in humans. Following infancy, ‘childhood’ spreads from 3 to 6 years of age when modern human weaned infants show all of their deciduous teeth in functional occlusion, although they cannot yet fully process an adult diet, due to their relatively small size and thin enamel5. In modern humans, the end of childhood is marked by the eruption of the permanent first molar (M1) at ~ 6 years of age1, and the attainment of adult brain size10.In this context, fossil teeth are especially valuable not only because their recovery enables comparing developmental stages between fossil hominins and modern humans but also because dental hard tissues record their own growth and maturation as well as stressful events experienced by the organism11. In the tooth microstructure, stress events may manifest as accentuated lines and remain non-specific12,13,14. A specific accentuated growth marking called the neonatal line is often identifiable in the enamel and dentine of the deciduous teeth and in the mesial cusps of the permanent first molar which start mineralizing in utero15, 16.During both infancy and childhood, protection and food provisioning by adults of the group (parents or others) is crucial for the child’s survival. Understanding the modality and time of emergence of this unique modern life history pattern is currently one of the most debated topics in human evolution9, 17, 18.GAR IVE (full name: MK 81 GAR IVE 0043) is a fragmentary right hemimandible with a partial mixed dentition, recovered in layer E at the Garba IV site of the Melka Kunture complex (Upper Awash, Ethiopia) (Fig. 1). This fossil was attributed to Homo erectus s.l.19, and its age at death was estimated to be 2–3.5 years20, 21. With the recent discovery of H. erectus s.l. remains in association with Acheulean and Oldowan artifacts at Gona (Afar, Ethiopia)22, GAR IVE is one of the rare early Homo remains recovered from the beginning of the Early Pleistocene, in the context of proper archaeological excavations providing contextual information on hominin activities23. GAR IVE was found with Oldowan industry and faunal remains in layer E, later dated to ~1.8 Ma (Fig. 1b–d)24,25,26,27,28. The deposits of Garba IV were dated and arranged in sequence in a thoroughly established chronostratigraphy26, 29, which was evaluated again during fieldwork in 2013 and 2017.Figure 1Garba IV site. (a) Location map of Melka Kunture and the Garba IV site (modified after https://upload.wikimedia.org/wikipedia/commons/2/24/Africa_satellite.jpg, https://commons.wikimedia.org/wiki/File:Awashrivermap.png, and https://data.humdata.org/dataset/ethiopia-elevation-model using QGIS software; https://qgis.org); (b) Plan of archaeological level E with position of GAR IVE (modified after Fig. 4A from Ref.27); (c) Stratigraphic log of Garba IV (Photoshop CS3; https://www.adobe-photoshop); (d) schematic cross-section of the archaeological levels (modified after Fig. 3B from Ref.27). The map of the Awash River (https://commons.wikimedia.org/wiki/File:Awashrivermap.png) is licensed under the Creative Commons Attribution-ShareAlike 3.0 license. The license terms can be found on the following link: http://creativecommons.org/licenses/by-sa/3.0/.Previous studies reported GAR IVE as being affected by a rare genetic disease amelogenesis imperfecta (AI)19, although they were only based on classical radiographs and SEM imaging of epoxy replicas of the tooth surface19, 30. Zanolli and colleagues worked on CT data, but did not re-evaluate this diagnosis21. Amelogenesis imperfecta encompasses a group of developmental conditions31 altering the enamel structure and making teeth fragile32. The main diagnostic criteria may involve a marked reduction in enamel thickness frequently concerning both deciduous and permanent teeth, enamel pits or linear hypoplasia not associated with any noticeable developmental defects in the other dental tissues, a brown colouration of the enamel, or taurodontism (enlargement of the pulp cavity associated with a lowering of the roof of the pulp chamber and root furcation level, giving the impression of a partial to complete fusion of the tooth roots)31,32,33,34,35,36. In some AI variants, enamel defects may be associated with other types of syndromes occurring in other parts of the human body (e.g., kidney diseases)32, 37, 38. In clinics, a differential diagnosis requires the knowledge of medical and family history of the patients, of their genetic profile, as well as of their conditions of development and life. Thus, AI patients mostly suffer from early tooth loss, severe embarrassment, eating difficulties and pain, requiring early and complex restorative treatments38.Genetic diseases are of particular interest as they happen to be transmitted along a lineage affecting multiple generations, and they may occur with variable frequencies, in the case of AI, ranging from 1:700 to 1:14 00038. According to Zilberman et al.19, 30, the presence of AI in GAR IVE would provide evidence of genetic continuity from early Homo to modern humans. Yet, this claim has to be considered with caution since the molecular mechanisms causing this pathology are complex and may involve several genes and combinations of genes39. Such a genetic continuity is unlikely to be so straightforward. Furthermore, Trinkaus40 has suggested that any variant of this pathology would have involved the need for enhanced maternal care to supply the young child with sufficient nutrients and permit its survival to some extent40.Given the putative consequences of this disease on the specimen’s morphology and the potential implications of living with AI during the Pleistocene, we aim to reassess the diagnosis of AI by analyzing tooth surface features and by exploring the fine internal structure of the bone and teeth using synchrotron imaging. We compare the GAR IVE teeth with teeth from modern human patients, some with documented forms of AI. Because taphonomic processes may affect the preservation of the specimens and the features related to the AI diagnosis, our study includes the analysis of the associated faunal remains and the geological context in which GAR IVE was recovered in order to establish if particular features are due to taphonomy or to developmental alterations induced by the AI. In spite of the taphonomic damages undergone by the dental tissues, we provide some novel fragmentary yet still valuable palaeobiological data on the dental development of this early Homo infant.After a brief description of the GAR IVE specimen, and especially of its dental development, we have compared the enamel features described as diagnostic of AI on GAR IVE to the enamel condition in teeth of modern-day AI patients. Then, we have explored the fine microstructure of the teeth and bone in light of taphonomic processes, recovered from the study of associated faunal remains, and the palaeoenvironmental reconstruction of the fossilisation context of GAR IVE.Description of GAR IVEThe GAR IVE right hemimandible (Fig. 2) preserves both erupted deciduous molars (LRdm1 and LRdm2). The LRdm1 is considerably worn (yet preserving a complete rim of enamel) and super-erupted above the occlusal level. The germ of the permanent first molar (LRM1) is partially exposed at the posterior part of the specimen, as the mandibular ramus is missing. Its crown does not present any abnormal morphological features; to note that its occlusal surface shows extensive wrinkling, yet this LRM1 looks similar to that of other fossil hominins (Fig. 3). The anterior portion of GAR IVE shows a greenstick fracture on its lingual side, suggesting this occurred peri-mortem, when the bone would have retained a certain level of collagen and moisture, thus still be fresh and elastic to some extent41. This fracture leaves the permanent lateral incisor (LRI2) visible and partially unprotected by its bony crypt. The germs of the permanent canine (LRC1), third and fourth premolars (LRP3 and LRP4) are unerupted and protected inside the mandibular bone. The bone and teeth present various kinds of surface alterations.Figure 23D models of the GAR IVE right hemi-mandible showing the mandibular bone (beige), the preserved deciduous molars (dm1 and dm2; pink) and the germs of the permanent teeth (I2: lateral incisor, C: canine, P3 and P4: first and second premolar, M1: first molar; blue). Superior, inferior, buccal, anterior, lingual and posterior views.Figure 3The occlusal surface of the LRM1 of GAR IVE (3D model) shows an amount of wrinkling comparable with that of other Plio-Pleistocene hominin molars such as the LRM2 of Omo 427 and the LLM3 of Omo 136-1 (*: flipped for illustrative purpose; both teeth were macro-photographed).Insights into GAR IVE’s dental developmentThe synchrotron µCT scans show a loss of contrast between bone, enamel and dentine, the enamel-dentine junction being barely visible in some places (Fig. 4). Any estimation of age at death based on dental growth increments and virtual histology techniques is thus precluded. At best, the maturation stage of the permanent tooth germs may provide insights into GAR IVE’s dental development (See Supplementary Text S1 for details). Following modern human standards42, the overall calcification stages in GAR IVE would correspond to 4.5 years of age (with individual scores per tooth going up to 7.5 years for the P3; see Supplementary Table S1). Using Kuykendall’s regression equation43, GAR IVE would be 3.13 [2.14–4.12] years in a chimpanzee-equivalent model. Several fossil hominins died at developmental stages similar to that of GAR IVE, although any comparison highlights some modularity in the relative advancement of each tooth type across taxa (See Supplementary Text S1). In terms of calcification stages, KNM-ER 812 (Paranthropus boisei) seems to be developmentally the closest to GAR IVE and has an estimated age at death of 2.5–3.0 years. KNM-ER 1820 (P. boisei) is slightly more advanced than GAR IVE for an age at death similar to KNM-ER 812 (2.5–3.1 years). Overall, the calcification of the GAR IVE permanent tooth germs is less advanced than that of Sts 24 (Australopithecus africanus).Figure 43D model of the GAR IVE specimen (a), and frontal (b, red), axial (c, blue) and sagittal (d, green) planes of section to illustrate the morphology of the hole (orange arrow) located in the area of the mental foramen. Its inner negative imprint shows a complex shape which is little compatible with the shape of a carnivore tooth (c). To note an additional convoluted shape represented in green in 3D (pink arrow).In spite of the taphonomic alteration of the dental tissues in GAR IVE, the synchrotron data revealed several accentuated lines in the LRP4 (Supplementary Text S1, Supplementary Fig. S13). These accentuated markings may correspond to multiple stressful events. However, it remains impossible to ascertain their cause which may result from non-specific stress events in the life of the child. It also remains unknown whether these accentuated markings may witness disruptions in the growth process of GAR IVE, or even in its health. Assuming the initiation time of P4 (~ 2 years) and that the EDJ crest where the virtual slice was recorded formed ~ 3 months after the dentine horn tip, these were added to the times of formation of
https://www.nature.com/articles/s41598-021-02462-1
Insights into the palaeobiology of an early Homo infant: multidisciplinary investigation of the GAR IVE hemi-mandible, Melka Kunture, Ethiopia